
|
Подскажите, пожалуйста, планируете ли вы возобновление программ высшего образования? Если да, есть ли какие-то примерные сроки? Спасибо! |
Инспектор
Вы можете этот курс.
Опубликован: 14.12.2009 | Уровень: специалист | Доступ: платный
Лекция 13:
Активные нейронные модели (А-сети)
А-сеть - модель, удовлетворяющая принципу устойчивого неравновесия
Экспериментальные модели нейронных сетей, строившиеся на основе гипотезы Емельянова-Ярославского, в принципе, обладают свойством активности. Однако предложенная Емельяновым-Ярославским схема работы нейронной сети, так же как и ее модификации, плохо согласуется с принципом устойчивого неравновесия. Возможна и другая схема. Неравновесным состоянием естественно считать состояние повышенной возбудимости. В соответствии с принципом устойчивого неравновесия достигаться максимальное неравновесие (высокая возбудимость) должно не в покое нервной клетки, а наоборот - за счет ее работы, т. е. возбуждений. Порог возбуждения неработающей нервной клетки должен не уменьшаться, а увеличиваться. При каждом срабатывании нейрона его порог возбуждения должен не увеличиваться, а уменьшаться. Оптимальное (неравновесное) состояние нейрона должно соответствовать его повышенной возбудимости, т. е. минимальному значению порога возбуждения.
Рассмотрим на качественном уровне соответствующий вариант активной сети. Экспериментально этот вариант не прорабатывался. Однако достоверность делающихся выводов и предположений во многом основана на опыте экспериментальной работы с рассмотренной модификацией схемы Емельянова-Ярославского, т. е. с моделью трехслойной сети.
В гипотетической модели (  -сети), так же как и ранее, предполагается, что нейрон - это пороговый элемент с временной и пространственной суммацией возбуждающих и тормозящих воздействий. Изменяющиеся параметры нейронов - это функциональное состояние, накопленный потенциал возбуждения, порог возбуждения и синаптические проводимости возбуждающих и тормозящих связей. Параметры, отражающие функциональное состояние
-сети), так же как и ранее, предполагается, что нейрон - это пороговый элемент с временной и пространственной суммацией возбуждающих и тормозящих воздействий. Изменяющиеся параметры нейронов - это функциональное состояние, накопленный потенциал возбуждения, порог возбуждения и синаптические проводимости возбуждающих и тормозящих связей. Параметры, отражающие функциональное состояние  и порог возбуждения
и порог возбуждения  нейрона, связаны прямой зависимостью
нейрона, связаны прямой зависимостью  . (В принципе, о параметре
. (В принципе, о параметре 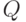 можно было бы не говорить, сводя все к порогу возбудимости, имеющему оптимальное значение и изменяющуюся величину.) Порог возбуждения нейрона увеличивается у несрабатывающего нейрона. При каждом срабатывании порог уменьшается на постоянную величину.
Так же изменяется и параметр
можно было бы не говорить, сводя все к порогу возбудимости, имеющему оптимальное значение и изменяющуюся величину.) Порог возбуждения нейрона увеличивается у несрабатывающего нейрона. При каждом срабатывании порог уменьшается на постоянную величину.
Так же изменяется и параметр  . Возбуждающий потенциал
. Возбуждающий потенциал  образуется алгебраическим суммированием приходящих по связям возбуждающих и тормозящих воздействий. Величины и знаки этих воздействий определяются величинами и знаками синаптических проводимостей связей
образуется алгебраическим суммированием приходящих по связям возбуждающих и тормозящих воздействий. Величины и знаки этих воздействий определяются величинами и знаками синаптических проводимостей связей  . Проводимости связей изменяются при прохождении по ним сигнала в зависимости от значения параметра нейрона, на который приходит сигнал. Между одновременно срабатывающими нейронами могут образовываться новые связи. Нейрон срабатывает при
. Проводимости связей изменяются при прохождении по ним сигнала в зависимости от значения параметра нейрона, на который приходит сигнал. Между одновременно срабатывающими нейронами могут образовываться новые связи. Нейрон срабатывает при  . При срабатывании нейрона возбуждающий потенциал становится равным нулю. При отсутствии воздействий на нейрон накопленный потенциал экспоненциально снижается. Свойства нейронов и связей между ними таковы, что оптимизация в сети возможна лишь при групповом взаимодействии (взаимосодействии) нейронов и их непрерывной или периодической активности.
. При срабатывании нейрона возбуждающий потенциал становится равным нулю. При отсутствии воздействий на нейрон накопленный потенциал экспоненциально снижается. Свойства нейронов и связей между ними таковы, что оптимизация в сети возможна лишь при групповом взаимодействии (взаимосодействии) нейронов и их непрерывной или периодической активности.
Начальные качественные предположения об основных общих свойствах нейронных механизмов -сети состоят в следующем:
- элементом памяти в -сети является нейронная структура (нейронный ансамбль) - группа нейронов, объединенных взаимовозбуждающими связями;
- эти структуры - ансамбли - формируются в сети при повторяющихся информационных воздействиях, приводящих к одновременному возбуждению группы нейронов;
- на ансамблях нейронов реализуются ассоциации по сходству - весь ансамбль может возбуждаться при приходе внешнего возбуждения только на часть его нейронов;
- между регулярно возбуждающимися последовательно во времени ансамблями устанавливаются возбуждающие связи в прямом направлении времени и тормозящие в обратном. На основе этих связей осуществляется предвидение (предактивация);
- система памяти (модель среды) имеет многоуровневое иерархическое строение, позволяющее формировать обобщения и укрупнения;
- каждый уровень модели среды реализуется в виде трехслойной сети с возвратным торможением;
- свойства нейрона выражаются зависимостями от некоторой переменной величины, соответствующей функциональному состоянию или критерию качества функционирования нейрона и зависящей в каждый момент от интенсивности его срабатываний в течение некоторого предшествовавшего периода (предыстории возбуждений);
- оптимизация по параметру "функциональное состояние" как отдельного нейрона, так и всей сети достигается за счет срабатываний нейронов при их групповом взаимосодействии;
- оптимизация функционального состояния нейронов при приходящих на сеть новых внешних информационных воздействиях достигается за счет структурной перестройки (память);
- при внешних мотивационных воздействиях оптимизация достигается только при снятии этих воздействий в результате целенаправленного поведения;
- решение любых задач в нейронной сети состоит в формировании и гашении очагов возбуждения, мешающих нормальному процессу нормализации общего функционального состояния сети.
Большинство приведенных предположений должны в основном рассматриваться как постулаты, которые определяют свойство активности нейронной сети, направленной на оптимизацию своего функционального состояния. Эти предположения в определенной степени пересекаются с общими исходными свойствами модели Емельянова-Ярославского.
Первично нейроны должны возбуждаться только при внешних активирующих информационных воздействиях. Одновременно возбуждающиеся нейроны образуют ансамбль. Связи между нейронами ансамбля создают синергическое возбуждающее взаимосодействие, обеспечивающее работу (возбуждение) ансамбля и при снятии внешних воздействий. Эта работа должна приводить к достижению оптимальных состояний нейронов ансамбля и их повышенной возбудимости. Заканчиваться работа ансамбля должна при поступлении на него тормозящих воздействий от других работающих ансамблей, а также от элементов, создающих возвратное торможение (схема с возвратным торможением описана выше). Последовательно возбуждающиеся ансамбли должны связываться в цепочки за счет возбуждающих связей в прямом направлении времени и тормозящих в обратном. Это достигается за счет принимаемых в модели свойств связей.
Проводимости связей изменяются при одновременном срабатывании связываемых нейронов. Проводимость возбуждающей связи от нейрона 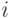 к нейрону
к нейрону 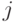 увеличивается, если
увеличивается, если  , и тем сильней, чем больше разность
, и тем сильней, чем больше разность  . Проводимость тормозящей связи от нейрона к нейрону
. Проводимость тормозящей связи от нейрона к нейрону 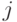 увеличивается, если
увеличивается, если  и тем сильней, чем больше разность
и тем сильней, чем больше разность  . По таким же правилам должны образовываться новые связи между нейронами. В результате будут образовываться ансамбли взаимовозбуждающихся нейронов. Кроме того, будут усиливаться возбуждающие связи, которые идут от ансамбля, начавшего работать раньше, к ансамблю, начавшему работать позже. Также будут усиливаться тормозящие связи, идущие от ансамбля, начавшего работать позже, к ансамблю, начавшему работать раньше.
. По таким же правилам должны образовываться новые связи между нейронами. В результате будут образовываться ансамбли взаимовозбуждающихся нейронов. Кроме того, будут усиливаться возбуждающие связи, которые идут от ансамбля, начавшего работать раньше, к ансамблю, начавшему работать позже. Также будут усиливаться тормозящие связи, идущие от ансамбля, начавшего работать позже, к ансамблю, начавшему работать раньше.
Если нейроны ансамбля не возбуждаются, то их пороги срабатывания увеличиваются. Одновременно увеличиваются (отклоняются от оптимума) значения параметров , определяющих функциональные состояния нейронов. Можно сказать, что в -сети при этом возникает потребность во внешнем информационном запускающем возбуждающем воздействии на ансамбли нейронов. Кроме того, возникает потребность в образовании ансамблей взаимосодействующих нейронов, достигающих оптимальных состояний только в процессе совместной работы и взаимных возбуждений. И наконец, возникает потребность в активации ансамблей от других ансамблей, работа которых при отсутствии внешних информационных воздействий выстраивается в регулярные временные последовательности (предвидение).
Есть еще одно принципиальное отличие от схемы Емельянова-Ярославского. Попытки понять, как устроены и работают нейронные механизмы мозга, должны основываться на том, что мозг развивался в процессе эволюции не для решения своих собственных задач, а для решения задач целого организма. Необходимо ответить на вопрос, как на основе неустойчивости и активности нейронных механизмов решаются не только внутренние, но и внешние функциональные задачи мозга. В первую очередь нужно ответить на вопрос, как осуществляется обучение, то есть построение в мозге активной иерархической модели проблемной среды, и как работает эта активная модель при решении задач восприятия и управления целенаправленным поведением живого организма, а также и при решении абстрактных задач.
Можно представить, что кроме кратковременных информационных возбуждений возможны и длительные возбуждения, мешающие режиму последовательной активации разных нейронных ансамблей, т. е. общей саморегуляции сети. Стабильные фиксированные очаги возбуждения могут соответствовать первичным и вторичным потребностям живого организма. Прекращение возбуждения должно происходить при удовлетворении потребности и снятии соответствующих возбуждающих воздействий. Так связываются поведенческие задачи с задачей оптимизации состояния нейронной сети.
Не менее интересен и вопрос о том, как в мозге возникают, фиксируются и умозрительно решаются задачи абстрактного мышления. В мозге при постановке и решении любых задач, т. е. не только задач поведения, но и умозрительных абстрактных задач, работает принцип доминанты А.А. Ухтомского. При моделировании мышления важно ответить на вопрос, как на нейронном уровне может происходить возникновение, фиксация и снятие доминант или возникновение, поддержание и гашение очагов возбуждения. Возникновение очагов возбуждения, мешающих общей равномерной оптимизации сети, и их гашение соответствует возникновению и решению поведенческих и абстрактных задач мышления, а также поиску информации, необходимой для достройки модели проблемной среды.
В гипотетической полной модели среднее значение параметра по всей сети должно быть "эмоциональной переменной", то есть должно учитываться в эмоциональном центре при выборе линии поведения наравне с другими эмоциональными переменными, формирующими первичные и вторичные мотивации.
И наконец, важно понять, как в нейронной модели может быть реализован полный отдых в режиме "сна". Этот режим должен приводить к равномерной нормализации функциональных состояний всех нейронов при отсутствии внешних информационных воздействий.
Помимо основных функциональных или рабочих свойств мозг проявляет и специальные свойства и функции, присущие только живому мозгу: это интуиция, озарение, или инсайт, творчество, эмоциональные реакции на музыку, абстрактную живопись (т. е. живопись без смысловой нагрузки), стихотворный ритм, красоту природных явлений. К этому перечню можно добавить еще любопытство, юмор, навязчивую идею, навязчивый мотив. Было бы хорошо, если бы нейронная модель объясняла и такие свойства. Не исключено, что именно активное устойчивое неравновесие нейронов и синергическая активность нейронной сети могут дать ключ к пониманию и этих свойств мозга. Некоторые интерпретации приводятся в дальнейшем изложении. Однако подробная проработка и тем более экспериментальная проверка логики работы активных нейронных сетей, соответствующих принципу устойчивого неравновесия, не проводились.
Владислав Нагорный
Лариса Парфенова
|
1) Можно ли экстерном получить второе высшее образование "Программная инженерия" ? 2) Трудоустраиваете ли Вы выпускников? 3) Можно ли с Вашим дипломом поступить в аспирантуру?
|