

|
Подскажите, пожалуйста, планируете ли вы возобновление программ высшего образования? Если да, есть ли какие-то примерные сроки? Спасибо! |
Инспектор
Вы можете этот курс.
Опубликован: 14.12.2009 | Уровень: специалист | Доступ: платный
Лекция 13:
Активные нейронные модели (А-сети)
Модель трехслойной активной сети с возвратным торможением
Рассмотрим экспериментальную модель [17], построенную на основе идеи о неустойчивости и активности нейронов. Отличие этой модели от общей схемы Емельянова-Ярославского состоит во введении сенсорного слоя и слоя элементов, обеспечивающих возвратное торможение.
Нейрон, как и любой целостный объект, может полностью характеризоваться набором переменных, их текущими значениями, зависимостями между переменными и зависимостями переменных от времени и внешних воздействий. Применительно к модели нейронной сети формальное задание всех этих зависимостей и аналитическое описание на этой основе работы системы в целом не представляется возможным. Поэтому все переменные в описываемой модели определяются пошаговыми рекуррентными соотношениями. Время дискретно, т. е. разбивается на такты. В каждом такте просчитываются и изменяются все переменные (параметры) всех нейронов и связей. Таким образом, строится програмная модель нейронной сети, рассматриваемой как физический объект.
Принимается (постулируется), что для моделирования нейронных механизмов мышления достаточны объекты двух типов: нервная клетка (нейрон) и нервная связь. Переменные, характеризующие нейрон, - это возбудимость, потенциал поляризации мембраны и функциональное состояние. Переменные, характеризующие связь, - это ее проводимость, топология, знак и временная задержка передаваемого сигнала. Сведение всех свойств нейрона к этому ограниченному множеству упрощает и делает практически возможным компьютерное моделирование. В то же время это сопряжено с опасностью игнорирования каких-то свойств нейрона как живой клетки, что в конечном счете может отразиться на принципиальных свойствах модели. Однако представляется, что этот риск не очень велик, если в модели сохраняются главные свойства живого - неустойчивость и активность.
Приведем общие свойства нейронов экспериментальной модели. Точные количественные характеристики широко варьировались при проведении экспериментов.
Функциональное состояние нервной клетки зависит от предыстории ее возбуждений и выражается в модели некоторым параметром  . Этот параметр имеет общее для всех нейронов оптимальное значение
. Этот параметр имеет общее для всех нейронов оптимальное значение  . Параметр дискретно увеличивается при каждом разряде нейрона и монотонно уменьшается во времени у несрабатывающего нейрона. Из этого следует, что для оптимизации функциональных состояний нервных клеток необходимы срабатывания (разряды) нейронов с некоторой оптимальной частотой. Нейрон может сработать при поступлении на его входы внешнего возбуждения. Достижение оптимального в среднем функционального состояния нейронов по всей сети достигается только при их взаимодействии - взаимном возбуждении или торможении.
. Параметр дискретно увеличивается при каждом разряде нейрона и монотонно уменьшается во времени у несрабатывающего нейрона. Из этого следует, что для оптимизации функциональных состояний нервных клеток необходимы срабатывания (разряды) нейронов с некоторой оптимальной частотой. Нейрон может сработать при поступлении на его входы внешнего возбуждения. Достижение оптимального в среднем функционального состояния нейронов по всей сети достигается только при их взаимодействии - взаимном возбуждении или торможении.
Нейрон представляет собой пороговый элемент с временной и пространственной алгебраической (т. е. с учетом знака) суммацией возбуждающих и тормозящих воздействий, переменной возбудимостью, рефрак-терностью и экзальтацией после срабатывания.
Условие срабатывания нейрона:  , где
, где 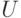 - порог срабатывания,
- порог срабатывания,  - накопленный потенциал.
- накопленный потенциал.
На нейрон может приходить любое количество возбуждающих и тормозящих связей от других нейронов или рецепторов. От нейрона отходит один аксон, который может разветвляться на любое количество возбуждающих и тормозных коллатералей, заканчивающихся на разных нейронах.
Накопленный на нейроне потенциал определяется алгебраической суммой пришедших на нейрон возбуждающих (+) и тормозящих (-) воздействий. Кроме того, накопленный потенциал у несрабатывающего нейрона экспоненциально уменьшается во времени:.

где:
 - значение накопленного потенциала в
- значение накопленного потенциала в  -м такте,
-м такте,
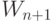 - значение накопленного потенциала в
- значение накопленного потенциала в  -м такте,
-м такте,
 - слагаемое, определяющее уменьшение накопленного потенциала во времени,
- слагаемое, определяющее уменьшение накопленного потенциала во времени,
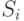 - величина синаптической проводимости
- величина синаптической проводимости 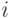 -й связи,
-й связи,
 - признак активности -й связи в -м такте.
- признак активности -й связи в -м такте.
После срабатывания нейрона накопленный на нем потенциал становится равным нулю.
Порог срабатывания имеет две составляющие - статическую и динамическую:  . B рассматриваемом промоделированном варианте статическая составляющая порога срабатывания монотонно снижается у несрабатывающего нейрона (возбудимость повышается) и увеличивается на малую дискретную величину при каждом срабатывании (возбудимость снижается). Статическая составляющая порога срабатывания связана прямой линейной зависимостью с параметром , т. е.
. B рассматриваемом промоделированном варианте статическая составляющая порога срабатывания монотонно снижается у несрабатывающего нейрона (возбудимость повышается) и увеличивается на малую дискретную величину при каждом срабатывании (возбудимость снижается). Статическая составляющая порога срабатывания связана прямой линейной зависимостью с параметром , т. е.  .
.
Динамическую составляющую порога срабатывания иллюстрирует рисунок 13.1.
Сразу после разряда порог срабатывания нейрона резко повышается, затем экспоненциально снижается до величины, меньшей чем начальное значение статической составляющей порога срабатывания (экзальтация), и затем повышается до величины, несколько превышающей начальное значение статической составляющей. Интервал  соответствует абсолютной рефрактерности, когда разряд нейрона невозможен при любых внешних воздействиях. Интервал
соответствует абсолютной рефрактерности, когда разряд нейрона невозможен при любых внешних воздействиях. Интервал  соответствует относительной рефрактерности, интервал
соответствует относительной рефрактерности, интервал  соответствует экзальтации.
соответствует экзальтации.
Связи между нейронами нужны для осуществления возбуждающих и тормозящих взаимодействий. От нейрона может отходить любое (в пределах технических ограничений) количество связей. Величина и знак передаваемого по связи потенциала определяются переменной величиной и знаком синаптической проводимости связи. Правила изменения синап-
тических проводимостей связей между нейронами наряду с правилами изменения порога возбудимости нейрона являются основой, определяющей работу нейронов и сети.
Синаптическая проводимость связи между -м и 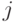 -м нейронами
-м нейронами  может "активно" изменяться при прохождении по этой связи сигнала. Приращение возможно только при условии срабатывания -ro нейрона. Если условий для "активного" изменения нет, синаптическая проводимость экспоненциально уменьшается, приближаясь к величине
может "активно" изменяться при прохождении по этой связи сигнала. Приращение возможно только при условии срабатывания -ro нейрона. Если условий для "активного" изменения нет, синаптическая проводимость экспоненциально уменьшается, приближаясь к величине  . Знак и величина "активного" изменения синаптической проводимости связи зависят от сочетаний значений величин
. Знак и величина "активного" изменения синаптической проводимости связи зависят от сочетаний значений величин 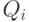 и
и  (функциональных состояний) пары нейронов в момент прохождения сигнала по связи между ними.
(функциональных состояний) пары нейронов в момент прохождения сигнала по связи между ними.
Общий качественный смысл зависимостей, определяющих приращения синаптических проводимостей, состоит в следующем. Величина положительного приращения возбуждающей синаптической проводимости связи от нейрона  к нейрону при прохождении по ней сигнала прямо зависит от разности
к нейрону при прохождении по ней сигнала прямо зависит от разности  . Кроме того, это приращение связано прямой зависимостью с разностью
. Кроме того, это приращение связано прямой зависимостью с разностью  . Наоборот, приращение проводимости тормозящего синапса увеличивается при увеличении в прямой зависимости от разности
. Наоборот, приращение проводимости тормозящего синапса увеличивается при увеличении в прямой зависимости от разности  .
.
Таким образом, при прохождении сигнала по связи ее синаптическая проводимость может увеличиваться, уменьшаться или оставаться неизменной в зависимости от сочетания значений  . При прохождении значения величины синаптической проводимости связи через нуль связь меняет знак, превращаясь из возбуждающей в тормозящую или наоборот. При моделировании рассматривался также вариант, в котором уменьшения значений синаптических проводимостей как возбуждающих, так и тормозящих связей ограничивались некоторой минимальной величиной
. При прохождении значения величины синаптической проводимости связи через нуль связь меняет знак, превращаясь из возбуждающей в тормозящую или наоборот. При моделировании рассматривался также вариант, в котором уменьшения значений синаптических проводимостей как возбуждающих, так и тормозящих связей ограничивались некоторой минимальной величиной  . В этом случае при
. В этом случае при  связь может уничтожаться, но менять знак не может.
связь может уничтожаться, но менять знак не может.
Изменения величин синаптических проводимостей связей между нейронами направлены на оптимизацию функционального состояния сети в целом и объединение во взаимовозбуждающую группу одновременно срабатывающих нейронов.
Все идущие от одного нейрона связи передают сигнал с одинаковой задержкой после срабатывания нейрона. (Вариант работы сети, в котором функциональную роль играют разные задержки сигналов, передаваемых по разным связям, мы рассмотрим в следующей лекции.)
Между одновременно возбуждающимися нейронами могут образовываться новые связи. Образование новых связей между нейронами происходит в соответствии с зависимостями, качественно совпадающими с зависимостями, по которым изменяются синаптические проводимости. Связи наиболее интенсивно образуются на те нейроны, величины которых, т. е. функциональные состояния, наиболее далеки от оптимальных значений. В области условий для образования новых связей нет.
В модели как постулат вводится принципиальное понятие - "функциональное состояние нервной клетки" и соответствующий ему параметр имеющий оптимальное значение . Можно вслед за Емельяновым-Ярославским давать этому параметру гипотетическую, не подкрепленную никакими экспериментами физиологическую интерпретацию, например, считая, что со значением параметра связана эффективность метаболизма (коэффициент полезного действия обменных процессов) нервной клетки.
Однако это не обязательно. Например, для целей моделирования достаточно считать, что средняя величина параметров нервных клеток является одной из физиологических констант живого организма, таких как, например, температура тела или артериальное давление. Удерживается эта величина в нужных пределах как средствами внутренней регуляции (взаимовозбуждение нейронов), так и средствами удовлетворения потребностей путем внешнего поведения (сон, отдых).
Описываемая нейронная сеть имеет три слоя. К исходной схеме Емельянова-Ярославского добавляется входной слой сенсорных элементов и второй слой нейронов. Нейроны второго слоя будем называть элементами. От нейронов первого слоя на элементы второго слоя образуются только возбуждающие связи. От элементов второго слоя на нейроны первого слоя образуются только тормозящие связи. В физиологии известны подобные схемы взаимодействия нейронов. Их название - структуры с возвратным торможением.
Рассматривались два варианта взаимодействия нейронов первого слоя и элементов второго слоя. В первом варианте тормозящая связь, идущая к нейрону первого слоя от элемента второго слоя, ничем не отличается от связей внутри первого слоя, и формируемый ею потенциал входит в алгебраическую сумму потенциалов нейрона. Во втором варианте тормозящая связь, идущая к нейрону первого слоя от элемента второго слоя, полностью запрещает срабатывание нейрона на время уменьшения создаваемого ею тормозного потенциала до некоторой величины. Все это время происходит независимая суммация всех других создаваемых на нейроне потенциалов.
Начальные условия эксперимента по формированию двухслойной первичной сети следующие: пороги возбуждения нейронов первого слоя и элементов второго слоя распределены случайно, равномерно, связей нет, внешние воздействия на сеть отсутствуют.
В переходном процессе формирования двухслойной первичной сети происходит распределение нейронов первого слоя по группам, объединенным торможением от общего элемента второго слоя, который преимущественно возбуждается нейронами этой группы. В установившемся режиме нейроны первого слоя чередуют срабатывания с интенсивностью, несколько превышающей оптимальную, и групповые синхронные срабатывания с периодом, равным времени спада тормозного потенциала. Элементы второго слоя срабатывают в фазе с групповыми срабатываниями нейронов первого слоя, чередуя эти срабатывания с паузами. При этом функциональные состояния нейронов первого слоя и элементов второго слоя, выражаемые величинами 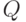 изменяются в некотором диапазоне вблизи
изменяются в некотором диапазоне вблизи  .
.
Таким образом, в процессе автоматического формирования двухслойной первичной нейронной сети образуется структура связей, разделяющая нейроны первого слоя на группы, которые объединены возбуждающими связями между нейронами внутри группы и возвратным торможением от общего для всех нейронов группы элемента второго слоя. Эти группы являются основой для образования вторичных нейронных сетей при внешних информационных воздействиях, то есть для запоминания этих воздействий.
В рассматриваемых в дальнейшем режимах работы сформировавшейся двухслойной нейронной сети на нейроны первого слоя подается "шумовое" возбуждающее воздействие, десинхронизирующее срабатывание нейронов. При этом срабатывание элемента второго слоя - случайное событие, имеющее вероятность, которая зависит от функционального состояния элемента и соответствующего функциональному состоянию порога возбудимости, т. е. в конечном счете от предыстории срабатываний.
Задача запоминания регулярных внешних воздействий решается по-разному в зависимости от того, каким образом связываются точки входного информационного слоя с нейронами первого слоя нейронной сети. В этом плане интересны два типа организации связей.
В первом случае имеется прямое соответствие: от каждой точки входного слоя, например, от рецептора, идет одна возбуждающая связь на "свой" нейрон первого слоя, и входное воздействие проектируется на нейронную сеть. В термин "проекция" не вкладывается геометрический смысл, так как распределение нейронов, соответствующих входному воздействию, не повторяет геометрических форм входного образа - нейроны выделяются равномерно по всей сети.
Во втором случае от каждой точки входного слоя отходит большое число возбуждающих и тормозящих случайных связей, оканчивающихся на разных нейронах сети. Входному образу до обучения соответствует равномерная передача возбуждающих и тормозящих воздействий на всю нейронную сеть.
Проводились два варианта экспериментов. В первом варианте от каждой точки входного информационного слоя имеется одна возбуждающая связь на соответствующий нейрон первого слоя сети. Точки входного слоя постоянно возбуждаются случайным образом. Входное воздействие подается на случайно выбранные точки входного слоя, синхронизирует возбуждение фиксированных точек, синхронизирует возбуждение соответствующих нейронов первого слоя и создает характерное распределение возбуждающего потенциала на элементах второго слоя, при котором вероятность сработать у них различна.
Элемент второго слоя, имеющий наибольшую вероятность возбуждения, начинает срабатывать в фазе с выделенной входным воздействием группой нейронов, в результате чего происходит перераспределение связей между слоями и образуется структурная группа - вторичная сеть. Выделенные информацией нейроны возбуждают один элемент второго слоя, через который подается возвратное синхронизирующее торможение на нейроны группы. Обучение происходит без подкрепления.
Описанный процесс возможен, если в сети между первым и вторым слоем имеется много лишних связей (заготовок), которые образовались на начальной стадии формирования первичной сети, а затем оказались избыточными для первичного разделения на группы, охваченные возвратным торможением, и уменьшили до Smin значения величин синапти-ческих проводимостей.
Во втором варианте обучение происходит с дифференцированным подкреплением. Необходимость подкрепления при обучении связана с тем, что в этом варианте от каждой точки входного слоя идет много случайных возбуждающих и тормозящих связей на нейроны первого слоя и при подаче любого входного возбуждения вся сеть возбуждается равномерно. Связи и величины синаптических проводимостей связей между точками входного слоя и нейронами первого слоя изменяются по зависимостям, сходным с зависимостями изменения связей в первом слое сети.
В процессе обучения периодически возбуждаются фиксированные, случайно выбранные точки входного слоя. Одновременно дается подкрепление - возбуждается один из элементов второго слоя сети. При этом происходит изменение синаптических проводимостей входных связей. От возбуждающихся точек входного слоя увеличиваются возбуждающие и уменьшаются тормозящие синаптические проводимости связей на нейроны первого слоя, объединенные возвратным торможением от элемента второго слоя, на который дается подкрепление. Синаптические проводимости возбуждающих связей, идущих от выделенных входным воздействием точек входного поля на все нейроны, которые не входят в подкрепляемую группу, уменьшаются, тормозящие увеличиваются, до тех пор, пока входное воздействие не перестает оказывать десинхронизирующего влияния на режим регулирования функциональных состояний в первичной сети.
Таким образом происходит привязка подкрепляемой группы первичной сети к выделенным информационным воздействием точкам входного слоя. После обучения поданное на вход "знакомое" кратковременное информационное воздействие запускает периодический процесс срабатывания одной группы - образованной вторичной сети, которая одновременно является группой первичной сети. Работа этой группы заканчивается за счет увеличения порогов срабатывания нейронов первого слоя и не оказывает десинхронизирующего влияния на работу других групп.
Во всех случаях обучение связано с перестройкой нейронной сети. Эта перестройка не должна искажать результатов предыдущего обучения, т. е. не должна затрагивать образованных при обучении вторичных нейронных сетей. В модели эта проблема может решаться формально - либо с помощью блокировок, либо при помощи направленного подкрепления. Физиологичность и того, и другого способа может вызывать сомнения.
Во всех вариантах обучения, если входной образ подается многократно с небольшими деформациями и искажениями, то после длительного процесса структурных изменений в сети сформированная входным воздействием структурная группа - вторичная сеть - включает только регулярно работающие нейроны первого слоя. Случайные искажения отбрасываются.
Если сеть обучена некоторому набору регулярных входных информационных воздействий, каждому из которых соответствует определенная вторичная сеть (ансамбль нейронов), то в системе возможны ассоциации по сходству: при подаче на сеть нового информационного воздействия, которое в общем случае может состоять из случайного набора точек входного поля, вероятность активации вторичных сетей, имеющих общие входные точки с этим входным воздействием, увеличивается. Чем больше общих входных точек, тем выше вероятность активации соответствующей вторичной сети, поскольку для запуска процесса срабатывания вторичной сети необходимо близкое по времени срабатывание входящих в нее нейронов первого слоя и входное воздействие синхронизирует срабатывания части этих нейронов.
После активации вторичной нейронной сети процесс некоторое время поддерживается сам за счет взаимных возбуждающих связей между входящими в группу нейронами первого слоя и возвратного синхронизирующего торможения. Таким же образом на вероятность активации вторичной сети влияет подача на вход искаженного знакомого информационного воздействия или его части.
На вероятность активации вторичной сети влияет не только число общих точек этой сети с входным воздействием, но и функциональные состояния, т. е. пороги возбудимости входящих во вторичную сеть нейронов первого уровня. Вследствие этого могут происходить отдаленные, т. е. не самые близкие по числу общих точек ассоциации.
Схема сети с возвратным торможением внешне похожа на схему классического трехслойного перцептрона. Так же, как в перцептроне, разделение пересекающихся на входном уровне информационных воздействий происходит на уровне групп нейронов первого слоя и связей, идущих к нейронам от сенсорных элементов. Принципиальное отличие - возвратное торможение и накапливающаяся неустойчивость нейронов. Последнее лежит в основе обучения и свойства активности. В описанной трехслойной сети возможно запоминание и спонтанное воспроизведение временных последовательностей, т. е. возможны ассоциации по смежности во времени. Однако экспериментальная реализация такого режима не проводилась.
Владислав Нагорный
Лариса Парфенова
|
1) Можно ли экстерном получить второе высшее образование "Программная инженерия" ? 2) Трудоустраиваете ли Вы выпускников? 3) Можно ли с Вашим дипломом поступить в аспирантуру?
|