
|
Подскажите, пожалуйста, планируете ли вы возобновление программ высшего образования? Если да, есть ли какие-то примерные сроки? Спасибо! |
Инспектор
Вы можете этот курс.
Опубликован: 14.12.2009 | Уровень: специалист | Доступ: платный
Лекция 15:
Основные проблемы, связанные с моделированием нейронных механизмов мозга
Умозрительное моделирование и свобода воли
Такой же важной, как проблема сознания, и трудноотделимой от него является проблема "умозрительного моделирования" и "свободы воли". Очевидно, что "умозрительное" моделирование в нейронной сети не должно сводиться к простому предвидению, основывающемуся на запоминании при обучении и воспроизведении временных последовательностей событий. Как ранее, так и в дальнейшем мы говорим о логической схеме сведения многопереборных многоэкстремальных задач к малопереборным одноэкстремальным, происходящего в мозгу человека или животного при решении задачи управления поведением в много-экстремальной среде. При этом приблизительно определяется структура используемой многоуровневой модели среды. Приблизительно определяется и общая логика происходящих в этой модели процессов.
Отчасти несколько забегая вперед, кратко можно сказать, что эти процессы выражаются в моделировании, эмоциональной оценке, сравнении и выборе среди разных вариантов переходов между текущей и целевой ситуациями разных уровней укрупнения в многоуровневой иерархической модели среды. Сначала оцениваются наиболее укрупненные и общие переходы, а затем по мере необходимости все более и более конкретизируемые. Кроме того, к умозрительному моделированию относятся и упоминавшиеся выше очень важные для понимания мышления процессы перцептивного и абстрактного конструирования в воображении.
Что заставляет мозг непрерывно в процессе поведения или абстрактного мышления упорядоченно и направленно выполнять всю эту работу и как она может реализоваться реальными и модельными нейронными механизмами? Так же как и при управлении сознанием, нейронный механизм управления "умозрительным" моделированием и сравнением вариантов должен формироваться автоматически на основе свойств нейронов, и это формирование должно подчиняться единому общему критерию оптимизации функциональных состояний нейронов и активной сети в целом. К этому критерию могут априорно (генетически) добавляться только какие-либо структурные характеристики сети, а также специфические (для конкретного случая) и неспецифические возбуждающие и тормозящие воздействия на сеть.
Это очень общее положение должно работать при построении любых конкретных нейронных механизмов. Для этого нужно придумать и экспериментально проверить необходимые свойства нейронов и сети.
Очень грубо и приближенно можно говорить о том, что при решении любой задачи в нейронной сети, в том числе и задачи абстрактного мышления, должны возникать очаги возбуждения - активированные нейронные ансамбли, соответствующие начальной и целевой ситуациям. Постоянная принудительная работа фиксированных ансамблей должна нарушать регулирование оптимальных состояний нейронов сети и стимулировать активацию цепочек ансамблей и переходов, связывающих эти ситуации. При моделировании необходимы привязанные к конкретным нейронным механизмам представления о формировании, поддержке и гашении очагов возбуждения.
В самом общем плане можно полагать, что для гашения очагов возбуждения и восстановления общей регуляции в сети должно быть необходимо физическое достижение целевых ситуаций, а при абстрактном мышлении активация очагов возбуждения одновременно с активацией нейронных ансамблей, соответствующих переходу между текущей и целевой ситуациями. При этом нужно отметить отдельную проблему - реализацию на нейронном уровне эмоциональных оценок переходов между ситуациями и сознательный направленный выбор на основе этих оценок наилучших вариантов переходов.
Осуществляемые в мозге в процессе мышления сознательный перебор, сравнение на основе эмоциональных оценок и фиксация лучшего варианта приводят к необходимости понимания не только на психологическом уровне, но и на уровне нейронных реальных и модельных механизмов еще одного феномена, который может быть назван "свобода воли". Я думаю, что проблема понимания и моделирования свободы воли не менее существенна и не менее сложна, чем проблема понимания и моделирования сознания.
Выше говорилось о проблемах моделирования воображения и управления целенаправленным сравнением воображаемых вариантов. Воображение может строиться на основе возбуждения существующих в модели проблемной среды нейронных ансамблей. Но этого недостаточно. При творческом мышлении используются перцептивное и когнитивное конструирование, т. е. воображаемое построение в сознании объектов, отсутствующих в модели среды, а может быть, отсутствующих и в самой среде. Для полного моделирования мышления необходимо конструктивное решение и этой проблемы.
Возникновение и гашение очагов возбуждения
Выше описывалось, как возникает очаг возбуждения в одноуровневой  -сети при подаче на нее стабильного информационного воздействия. Описывалось, как при этом активируется или формируется вторичная нейронная сеть, т. е. ансамбль нейронов, являющийся элементом памяти. При конкретной разработке многоуровневой нейронной модели необходимо будет определить логику и механизмы возникновения и гашения очагов возбуждения на всех уровнях нейронной сети.
-сети при подаче на нее стабильного информационного воздействия. Описывалось, как при этом активируется или формируется вторичная нейронная сеть, т. е. ансамбль нейронов, являющийся элементом памяти. При конкретной разработке многоуровневой нейронной модели необходимо будет определить логику и механизмы возникновения и гашения очагов возбуждения на всех уровнях нейронной сети.
Очаг возбуждения - это постоянно либо часто работающий (возбужденный) отдельный нейронный ансамбль или группа нейронных ансамблей. На возникновении и гашении очагов возбуждения в гипотетической полной нейронной -сети строятся все представления о ее работе при восприятии, поведении и мышлении. Возможно, что подобным же образом работают и реальные нейронные механизмы.
В
"Активные нейронные модели (А-сети)"
говорилось об экспериментах, когда в -сети создавался очаг возбуждения путем подачи на часть нейронов внешних возбуждающих воздействий. Очаг возбуждения нарушал работу сети, что приводило к образованию вторичной нейронной сети - элемента памяти. В более или менее общем виде можно представить и промоделировать появление в нейронной сети очагов возбуждения при возникновении первичных генетически определенных физиологических потребностей.
Первичным потребностям соответствуют первичные доминанты и отображающие их нейронные ансамбли. Как уже говорилось, доминанта, по А. Ухтомскому, - это стабильно сохраняющийся очаг возбуждения. При наличии потребностей происходит возбуждение соответствующих нейронных ансамблей с интенсивностями, определяемыми величинами потребностей. В соответствии с принципом доминанты А.А. Ухтомского на основе сравнения потребностей и с учетом текущей внешней ситуации формируется и фиксируется главная поведенческая доминанта и тормозятся остальные. Одновременно передается возбуждение на те нейронные структуры, которые должны участвовать в решении поведенческой задачи и снятии доминанты, т. е. в гашении очага возбуждения.
В соответствии с рассмотренными выше гипотезами доминанта возникает при оптимизационном решении поведенческой задачи по критерию maxT. От главной доминанты, т. е. очагов возбуждения самого верхнего уровня обобщения и укрупнения модели проблемной среды, на более низкие уровни распространяется конус, или "дерево", возбуждений. Одновременно передается торможение на нейронные ансамбли, соответствующие альтернативным переходам. На каждом уровне иерархической модели среды выбирается и фиксируется своя временная производная доминанта, соответствующая выбранному на этом уровне действию (переходу).
Доминанта (очаг возбуждения) гасится при реальном выполнении соответствующего перехода. Наибольшую трудность при нейронном моделировании таких процессов будет, по-видимому, представлять реализация перебора вариантов поведения, т. е. реализация переходов на всех уровнях нейронной модели проблемной среды, сравнение вариантов по их эмоциональной оценке и фиксация выбранного варианта в виде длительных и временных очагов возбуждения.
Моделированием процесса возникновения и гашения очагов возбуждения при формировании доминант, соответствующих генетически заданным первичным потребностям, проблема далеко не исчерпывается.
В соответствии с рассмотренной логикой очаги возбуждения должны возникать и гаситься в нейронной сети также и при возникновении и удовлетворении вторичных потребностей, при постановке и решении абстрактных, в том числе творческих, задач, при достройке и коррекции модели проблемной среды.
Четких представлений о том, как все это происходит в реальных или моделируемых нейронных сетях, нет. Нет также конструктивных гипотез, т. е. представлений на уровне нейронных механизмов, о том, как возникают, поддерживаются и изменяются постоянные вторичные потребности, т. е. потребности, не заданные генетически. То же самое относится и к временным вторичным потребностям, связанным с постановкой и решением абстрактных, в том числе и творческих, задач мышления.
Любые потребности сводятся в конечном счете к потребности в гашении очагов возбуждения. В наиболее общем плане мышление основывается на формировании и гашении очагов возбуждения. Очаги возбуждения наряду с потребностью поддержания оптимальных функциональных состояний нейронных элементов являются причиной и источником активности мозга. Поэтому можно сказать, что мыслящая система должна не только уметь, но и хотеть мыслить.
Можно сказать, что в нейронной сети, состоящей из активных неустойчивых элементов, постоянно возникают внутренние потребности в оптимизации своего функционального состояния. Никаких других потребностей в нейронной сети нет. Возникновение потребностей по оптимизации состояния связано с формированием в сети стабильных очагов возбуждения, приводящих к неравномерному возбуждению нейронов сети, что влечет за собой ухудшение функционального состояния отдельных нейронов и сети в целом. Удовлетворение таких потребностей достигается за счет гашения очагов возбуждения.
Эти свойства нейронов и нейронной сети должны быть заданы генетически. Кроме того, генетически заданы внешние по отношению к нейронной сети первичные физиологические потребности и их отображение в нейронной сети в виде стабильно поддерживаемых первичных очагов возбуждения. Гасятся такие очаги возбуждения только при удовлетворении первичных потребностей. Как возникают вторичные очаги возбуждения, связанные с вторичными потребностями, остается вопросом как на физиологическом уровне, так и на уровне моделирования. Точно так же вопросом остается и то, как работают реальные и модельные нейронные механизмы при возникновении и гашении вторичных очагов возбуждения в случае построения, коррекции или достройки модели среды.
Иногда говорят о том, что одной из генетически заданных первичных физиологических потребностей является получение информации. Это неверно. В описанном выше эксперименте Ротенберга и Аршавского в мозгу крысы при постоянном наблюдении приоткрытой двери в незнакомое темное помещение строился незаконченный фрагмент модели проблемной среды и формировался вторичный очаг возбуждения, погасить который можно, только исследовав это помещение и достроив фрагмент модели.
Таким образом, получение информации само по себе (любопытство) потребностью не является. Потребность же состоит в необходимости оптимизации функционального состояния нейронной сети при его ухудшении за счет появления очагов возбуждения. Вторичные очаги возбуждения гасятся, в частности, при активном и целенаправленном получении информации, необходимой для достройки целостной модели среды. Целенаправленность состоит в том, что по информации, имеющейся в существующей модели среды, строятся и затем проверяются гипотезы о свойствах исследуемого неизвестного объекта или исследуемой новой ситуации. Причем не о свойствах вообще, а о свойствах, имеющих смысл в отношении поведенческих задач, определяемых как первичными, так и вторичными потребностями.
Получение и запоминание признаков, отношений и метрических характеристик
Эта проблема, казалось бы, значительно более низкого уровня и может быть сопряжена при моделировании с очень серьезными затруднениями. Выше отмечалось, что входное поле любого, в том числе и первого - самого низкого, уровня -сети, не отождествляется с рецепторным, например, с сетчаткой глаза, хотя в частном случае при растровом распознавании такое отождествление возможно. В общем случае "вертикальные" входы какого-то уровня иерархии возбуждаются при активации информационных элементов другого уровня иерархии: например, на уровень слов должны подаваться входные воздействия с уровня букв или с уровня фонем. Части объединяются в целое не просто по принципу "И". Должны также формироваться "горизонтальные" связи между смысловыми элементами одного уровня. Такими связями, в частности, должны задаваться отношения и метрические характеристики.
Формирование элементарных смысловых единиц типа "признак", "отношение" или "метрическая характеристика" не только в -сети, но и ни в какой другой нейронной модели не рассматривается. Так, в ПРНС на вход сразу поступают признаки, полученные где-то и как-то вне нейронной модели. Таким образом, уже на самом нижнем уровне при входе в нейронную модель возникает проблема формирования в нейронной сети признаков. Проблема формирования отношений и метрических характеристик будет возникать на всех уровнях нейронной модели.
В зрительном анализаторе животных имеются рецептивные нейронные поля, выделяющие простые первичные признаки, такие как контуры, линии, направления линий, движения, грубая оценка размера объекта (большой, маленький). Эти и подобные простейшие преобразования входной информации в нейронной сети можно моделировать, но их явно недостаточно. В анализаторах входной информации животных и человека, по-видимому, происходит последовательная, усложняющаяся, многоуровневая обработка, направленная на формирование все более сложных характеристик воспринимаемых объектов и ситуаций.
Как строить на нейронах функции формирования в общем случае сложных произвольных признаков, отношений и метрических характеристик, неизвестно.
Неизвестно также, как на нейронах или нейронных ансамблях выделять, передавать, запоминать и сравнивать количественные характеристики. В частности, это относится к количественной эмоциональной оценке, необходимой для сравнения вариантов поведения. В экспериментах, о которых говорилось в "Физиологические интерпретации формальной модели поведения" , собака выбирала вариант поведения в соответствии с формулой выбора:

где:  - мотивация,
- мотивация,
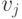 - скорость удовлетворения соответствующей потребности.
- скорость удовлетворения соответствующей потребности.
Собака, конечно, делала приблизительные оценки, но тем не менее в мозгу собаки каким-то образом на нейронном уровне при обучении запоминались ожидаемые величины скоростей удовлетворения потребностей, оценивались величины соответствующих мотиваций и в процессе сравнения вариантов определялись какие-то величины, коррелирующие с произведением  . (вряд ли собака производила точное умножение).
. (вряд ли собака производила точное умножение).
Таким образом, еще раз подчеркнем, что в мозгу определяются и запоминаются величины (количества), а также производятся операции с этими величинами. Простейшая из необходимых операций с количествами - это операция сравнения (больше-меньше). При построении моделей необходимо понять, как это делается нейронными механизмами мозга.
Владислав Нагорный
Лариса Парфенова
|
1) Можно ли экстерном получить второе высшее образование "Программная инженерия" ? 2) Трудоустраиваете ли Вы выпускников? 3) Можно ли с Вашим дипломом поступить в аспирантуру?
|